Nine years ago, Alan Weisman posed the scenario “The World Without Us.” The
premise was that, all of a sudden, people disappear entirely from the world. "What happens next?” The rest of the book described
the slow decay of buildings, roads, bridges, and other infrastructure, and the
gradual encroachment of wildlife on formerly human dominated landscapes. The
same scenario has been postulated in various movies, including Twelve Monkeys,
where humans dwelling underground send out hazmat-suited convicts to collect
biological samples from the surface in hopes of a cure for the devastating disease that destroyed most of humanity. The images of lions on buildings and bears in streets can seem as jarring – ok maybe not quite as jarring – as the
Nazi symbols on American icons in the Amazon.com adaptation of Philip K. Dick’s
The Man in the High Castle.
 |
| Twelve Monkeys |
The premise of this blog post is related – but even more dramatic
– what if evolution stopped – RIGHT NOW? What would happen? The context for
this question is rooted in my recent uncertainty, described in a paper and
my book, about how eco-evolutionary dynamics might be – mostly – cryptic. That
is, whereas most biologists seek to study eco-evolutionary dynamics by asking how
evolutionary CHANGE drives ecological CHANGE (or vice versa), contemporary evolution
might mostly counteract change. A classic example is encapsulated by so-called Red Queen
Dynamics, where it takes all the running one can do just to stay
in the same place. More specifically, everything is evolving all around you (as a species) and
so, if you don’t evolve too, you will become maladapted for other
players in the environment, which will cause you to go extinct. The same idea
is embodied – at least in the broad-sense – in the concept of evolutionary rescue,
whereby populations would go extinct were it not for their continual evolution
rescuing them from environmental change.
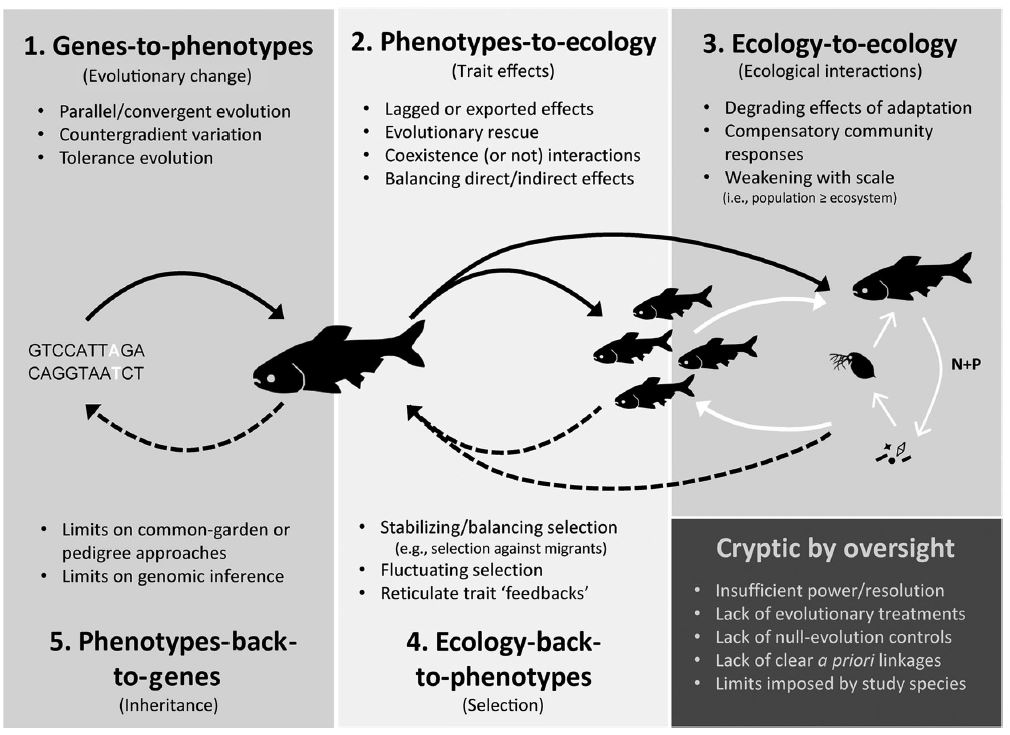 |
| From Kinnison et al. (2015) |
So how does one study cryptic eco-evolutionary dynamics? The current
gold standard is to have treatments where a species can evolve and other
treatments where they cannot, with ecological dynamics contrasted between the two cases.
The classic example of this approach is that implemented by Hairston, Ellner, Fussmann,
Yoshida, Jones, Becks, and others that use chemostats to compare predator-prey
dynamics between treatments where the prey (phytoplankton) can evolve and
treatments where they cannot. This evolution versus no-evolution treatment was achieved by the former having clonal variation present (so
selection could drive changes in clone frequencies) and the
latter having only a single clone (so selection cannot drive changes – unless
new mutations occur). These experiments revealed dramatic effects of evolution
on predator-prey cycles, and a number of conceptually similar studies by other investigators have yielded similar results (the figure below is from my book).

One limitation of these experiments is that the evolution
versus no-evolution treatments are confounded with variation versus
no-variation treatments. That is, ecological differences between the treatments could
partly reflect the effects of evolution and partly the effects of variation
independent of its evolution. An alternative approach is a replacement study,
where the same variation is present in both treatments and, although both might initially respond to selection, genotypes in the no-evolution treatment are
continually removed (perhaps each generation) by the experimenter and replaced
with the original variation. In this case, you still have an evolution versus
no-evolution treatment, but both have variation manifest as multiple genotypes – at least at the
outset.
All of these studies – and others like them – impose treatments on a single focal species, and so the question is “what effect
does the evolution of ONE species have on populations, communities, and ecosystems?” Estimates of the
effect of evolution of one species on ecological variables in nature,
regardless of the method, are then compared to non-evolutionary effects
of abiotic drivers, with a common driver being variation in rainfall. These
comparisons of "ecology" to "evolution" (pioneered by Hairston Jr. et al. 2005) generally find that the evolution of one species can have as large an
effect on community and ecosystem parameters as can an important abiotic driver, which
is remarkable given how important those abiotic drivers (temperature, rain,
nutrients, etc.) are known to be for ecological dynamics (the figure below is from my book).

A more beguiling question is “how important is ALL
evolution in a community?” Imagine an experiment could be designed to quantify the total effect
of evolution of all species in a community on community and ecosystem
parameters. How big would this effect be? Would it explain 1% of the ecological variation?
10%? 90%? Presumably, evolutionary effects of the whole community won’t be a
simple summation of the evolutionary effects of each of the component species.
I say this mainly because studies conducted thus far show that single species –
albeit often “keystone” or “foundation” species – can have very large effects on
ecological variables. A simple summation of these effects across multiple species
would, very soon, leave no variation left to explain. Hence, the evolution of
one species is presumably offset to some extent by the evolution of other
species when it comes to emergent properties of the community and ecosystem.
It is presumably impossible to have a real experiment with evolution and
no-evolution treatments at the entire community level in natural(ish) systems. We must therefore address the question (What would happen if all evolution ceased
RIGHT NOW?) as a thought experiment.
I submit that the outcome of a world without evolution experiment would be:
- Within hours to days, the microbial community at every place in the world will shift dramatically. The vast majority of species will go extinct locally and a few will become incredibly abundant - at least in the short term.
- Within days to weeks, many plants and animals that interact with microbes (and what organisms don’t?) will show reductions in growth and reproduction. Of course, some benefits will also initially accrue as – all of a sudden – chemotherapy, antibiotics, pesticides, and herbicides become more effective. The main point is that the performance of many plants and animals will begin to shift within a week.
- Within months, the relative abundance and biomass of plants and animals will shift dramatically as a result of these effects changing microbial communities and their influence on animal and plant performance.
- Within years, many animals and plants will go extinct. Most of these will go extinct because the shorter-lived organisms on which they depend will have non-evolved themselves into extinction.
- Within decades, the cascading effects of species extinction will mean than most animals and plants will go extinct, as will the microbes that depend on them. The few species that linger will be those that are very long lived and that have resting eggs or stages.
- Within centuries, all life will be gone. Except tardigrades, presumably.

The above sequence, which I think is inevitable, suggests several
important points.
1. Microbial diversity – and its evolution – is probably the
fundamentally irreducible underpinning of all ecological systems.
2. Investigators need to find a way to study the
eco-evolutionary STABILITY, as opposed to just DYNAMICS.
3. Evolution is by far the most important force shaping the
resistance, resilience, stability, diversity, and services of our communities
and ecosystems.
Fortunately, evolution is here to stay!